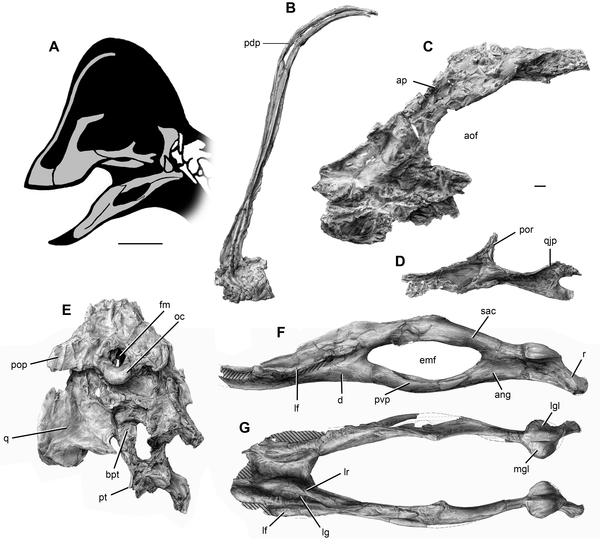
 |
| Elementi cranici riferiti ad Anzu (da Lamanna et al. 2014). |
Caenagnathidae è uno dei cladi più controversi di
Coelurosauria.
La sua composizione e diversità, in particolare per le forme
nordamericane, è complicata dal fatto che numerose specie sono state definite
su resti frammentari che non possono essere confrontati direttamente con altre
specie dello stesso clade. Ciò fa nascere il sospetto lecito che alcune specie,
in particolare quelle definite su resti non-confrontabili tra loro e
provenienti dalle medesime formazioni, siano di fatto sinonimi tra loro.
Ad esempio, le due specie di Caenagnathus sono basate
esclusivamente su mandibole. Idem per Caenagnathasia. Chirostenotes
invece non comprende resti di mandibole. Macrophalangia è basato su
resti degli arti posteriori. Elmisaurus è istituito su resti di
metapodiali. Hagryphus è basato su una mano. Epichirostenotes è
basato su resti sia craniali che postcraniali, ma manca di mandibola, mani e
piede, quindi non è comparabile direttamente con buona parte degli altri
caenagnathidi citati.
Da oltre un decennio era nota la scoperta di resti
moderatamente completi, appartenenti a più esemplari di un grande
caenagnathidae dalla Formazione Hell Creek. Questi resti sono stati pubblicati
in questi giorni (Lamanna et al. 2014) e sono riferiti ad un nuovo taxon, Anzu
wyliei.
Anzu presenta una combinazione di caratteri presenti
in altri caenagnathoidi, assieme a caratteri inattesi. Il premascellare
partecipa ad una alta cresta simile a quella dell'oviraptoride Rinchenia,
e si proietta posterodorsalmente oltre il livello della finestra antorbitale.
La fossa antorbitale ed i recessi relativi sono assenti. Il mascellare è
robusto e convesso anteroventralmente. Gli pterigoidi sono fusi medialmente. La
mandibola è molto simile a quella di Caenagnathus. La regione
retroarticolare è invece simile a Gigantoraptor. Come questo ultimo,
l'omero ha una torsione laterale in vista anteriore. Gli ungueali della mano
sono simili a Chirostenotes ed Elmisaurus. Il pube ha una fossa
prossimomediale, come Microvenator, Epichirostenotes e Nomingia.
L'ischio è molto simile a quelli di Chirostenotes e Epichirostenotes.
Purtroppo, il piede è quasi del tutto mancante, e ciò impedisce un confronto
con gli altri caenagnathidi.
Non ci sono dubbi che Anzu sia filogeneticamente
affine ai taxa menzionati. Immesso in una versione ridotta di Megamatrice
focalizzata sugli oviraptorosauri (ingroup a sua volta ottenuto dall'analisi
dell'intera matrice), Anzu (ed i nuovi caratteri presenti nello studio
di Lamanna et al. 2014) genera una topologia in parte conforme con quella di
Lamanna et al. (2014). Il risultato più interessante – e difforme da quello in
Lamanna et al. (2014) – è la risoluzione di un “grado caenagnathide” che forma
una serie parafiletica pettinata che forma la radice dei caenagnathoidi. In
particolare, Nomingia risulta la forma più basale, seguita da Epichirostenotes.
All'interno di Caenagnathidae abbiamo un ramo “elmisaurino” ed uno
“caenagnathino”. La relativa debolezza dei nodi non stupisce, dato che, come
accennato ad inizio post, la maggioranza dei caenagnathidi è basato su resti
non comparabili tra loro. Anche se Anzu porta una ventata di
informazioni per collegare i differenti morfotipi mandibolari e appendicolari
dei caenagnathidi, occorrono ulteriori esemplari relativamente completi per
irrobustire il quadro.

Bibliografia:
Lamanna
MC, Sues H-D, Schachner ER, Lyson TR (2014) A New Large-Bodied
Oviraptorosaurian Theropod Dinosaur from the Latest Cretaceous of Western North
America. PLoS ONE 9(3): e92022. doi:10.1371/journal.pone.0092022


